This is sort of old news but it is sometimes still worth mentioning. Back in March of this year there was another story that came out of a person who got cosmetic limb lengthening surgery. Unlike most of the people you hear about, this person was a girl.
While it is not very often we find girls who decide to go through with the surgery, there have been enough cases of girls who do it for the prospect of becoming a model. This girl fits that type of case study.
A 5′ 4″ tall aspiring model Alexandra Transer got the surgery done for a supposed 6,000 pounds and she has already gotten 6 cms out of it. She comes from a family of people who are below statured. Her mother is 5′ 2″ and her father is 5′ 5″. She chose not to go to University but work for her father in Engineering and saved up the 6,000 UK pounds to get the surgery.
She had wanted to be a model for most of her life, but when she contacted a modeling agency, they told her that she was below the height requirement and that the only way for her to get to that strict modeling height requirement was to extend her legs. She decided to do just that and went through with it, which we found very impressive and brave.
This first surgery she has decided to do is one of a total of 3 planned. There is supposed to be 2 more surgeries she will go through which will eventually put her at 6 feet. Her estimated time frame is that the 3rd surgery will be done by 2018. She will be in her 30s by then and may not get any type of modeling work but she did say that she plans to dye her hair blonde and then feel like a model being tall and blonde.
It seems that maybe what she really wants is to feel beautiful. On this website, we don’t judge. Vanity is something we all go through.
Some new information we learned is that she is actually from Russia herself, Volgograd. For anyone familiar with the LLS community, Volgograd is actually really famous because of the Centre of Anthropometrical (Orthopaedical) Cosmetology and Correction. Refer to Dr. Alexander Barinov and Dr. Viktor Shatov, of the center RUCOSM. Dr. Alexander Barinov is famous for being a protege and student to the more legendary Dr. Yegorov.
She mentioned that a former Soviet Athlete, a Valery Brumel, got this type of surgery done on them in 1968. Why did this olympic athlete get it done back in the 60s? We don’t know about that. I personally suspect that one of the reasons she was willing to go through with the surgery unlike so many other people who wish to be taller but are too afraid of surgery is because she comes from Russia, and specifically her home town has one of the most popular and recognizable centers for height increase surgery in the world. I am sure her parents at first was very reserved in her desires but they probably had a better understanding for why she would go through with it than other parents. Since they are from Russia originally, going a center like RUCOSM would not be as difficult of a transition as other people from other countries who might know any of the local Russian customs or language. Maybe for this girl, going to get limb lengthening surgery is as simple as driving the 3 miles to one’s local gym center to exercise.
Based on current conversion rates, it seems like she paid about $9,400 USD to get the first surgery.
We here have done our own level of research on LLS, and the cost of it. Most people have said that the cost of getting LLS is usually about $50,000 with a trip to one of the more well known, reputable surgeons costing upwards of $100,000 with everything covered, for maybe 10 cm or 4 Inches of increase in the leg bones. The lowest cost I personally have heard of was around $10,000 for the cost of surgery from a former Chinese surgeon. The reduced price of getting one’s leg extended seems to suggest that the cosmetic procedure may be becoming much more prevalant. That may be a good thing for some people who really want to go through with it and are looking for a cheaper alternative.
When we looked at the cost for LLS back in 2011, it seems that the cost was around 12,000 Euros, at least in the RUCOSM center, in Volgograd. Our source was from the Short Support Group website. The 12,000 euros cost was supposed to cover pretty much everything, including a 3 month stay. Of course, from the RUCOSM website, they state that for each surgery, they can only increase one’s bone length by 7-9 cm maximum. This is why Alexandra needed 3 total surgeries, to gain 8 inches or 20 cms of height to reach her desired goal of 6 feet tall from her original 5′ 4″. For her, she was very happy with the results, and there were no complications.
How much does the height increasing procedure cost? – Dr.Yegorov works with different clinics, both state and private. The total cost of the procedure including all necessary expenses is from $15,000 to $30,000 US dollars depending on the clinic and the accommodation and service conditions.
We would say that $10,000 USD is a little unrealistic of cost. A more reasonable cost would be probably $25,000 and that would just be for 7-8 cms of increase, assuming that no complications are developed. Of course those will be 2011 prices. Since it is 2015, We should expect maybe $30-35 K in fees in total. How Alexandra was able to just pay $10,000 we are not sure. Maybe she got her surgery done in the UK, and not Russia. Maybe instead of paying the thousands of dollars in fee for staying at the hospital facility to recuperate, she was able to have someone take her to her home in Volgograd (which might just be a few blocks away) during that time, allowing her to save thousands in hospital fees.
How does bone know how to be the proper shape and size for development? Can we manipulate this to grow taller?
I received this email from the author regarding how distraction osteogenesis would affect how bone manipulates growth in regards to maintaining placement of superstructures:
“It is indeed an interesting question as it challenges the system with an unnatural manipulation – i.e. interstitial growth.
The simple answer is: we haven’t tried, so I can’t say for sure.
If the relative locations of ligament and tendon insertions are what you are interested in, then previous works show that the periosteum is involved in regulation of their positions (see list below). Moreover, if the balance between proximal and distal growth rates is what you are interested in, then other works show that cross-sectional cutting and stripping of the periosteum can cause temporal acceleration in overall growth rate of the bone (also in humans, if I remember correctly), followed by a potential change in proximal to distal growth balance (I don’t think that these works test how these influence the positioning of superstructures in the bone; see list below). Therefore, if the operation you are applying includes anchoring of the periosteum to the bone or its cutting and stripping, this is something that may influence the scaling of the bones.”
“One of the major challenges that developing organs face is scaling, that is, the adjustment of physical proportions during the massive increase in size. Although organ scaling is fundamental for development and function, little is known about the mechanisms that regulate it. Bone superstructures are projections that typically serve for tendon and ligament insertion or articulation and, therefore, their position along the bone is crucial for musculoskeletal functionality. As bones are rigid structures that elongate only from their ends, it is unclear how superstructure positions are regulated during growth to end up in the right locations. Here, we document the process of longitudinal scaling in developing mouse long bones and uncover the mechanism that regulates it. To that end, we performed a computational analysis of hundreds of three-dimensional micro-CT images, using a newly developed method for recovering the morphogenetic sequence of developing bones. the relative position of all superstructures along the bone is highly preserved during more than a 5-fold increase in length, indicating isometric scaling. It has been suggested that during development, bone superstructures are continuously reconstructed and relocated along the shaft, a process known as drift. most superstructures did not drift at all. Instead, we identified a novel mechanism for bone scaling, whereby each bone exhibits a specific and unique balance between proximal and distal growth rates, which accurately maintains the relative position of its superstructures. Moreover, we show mathematically that this mechanism minimizes the cumulative drift of all superstructures, thereby optimizing the scaling process. [There’s] a general mechanism for the scaling of developing bones. More broadly, these findings suggest an evolutionary mechanism that facilitates variability in bone morphology by controlling the activity of individual epiphyseal plates.”
If we can trick the bone into thinking it’s drifting maybe we can convince it to grow to maintain the position of the superstructure. For example, dislocating the bone or similar means.
Although the molecular mechanisms regulating each growth plate for different bones are similar the bones still have different elongation rates.
“superstructures, known as bone ridges, tuberosities, condyles, etc., are necessary for the attachment of tendons and ligament as well as for articulation. To perform these functions they are located at specific positions along the bone. Bone superstructures emerge during early skeletogenesis . During growth, bones elongate extensively by advancement of the two growth plates away from the superstructures. It is therefore expected that during elongation, superstructures would remain at their original position near the center of the bone. Nevertheless, the end result is proper spreading of superstructures along the mature bone, which clearly implies the existence of a morphogenetic mechanism that corrects their locations.”
It’d be interesting to see what happens to bone superstructures during distraction osteogenesis.
“An ossified bone is a rigid object and so are the superstructures protruding from it, implying that they cannot be relocated by means of cell migration or proliferation. Therefore, any scaling mechanism must be adapted to overcome these physical restrictions.”<-So we have to make the bone less rigid.
“forelimb bones tend to grow away from the elbow joint, whereas bones in hind limbs tend to grow toward the knee joint.”
” Because the periosteal sheath is stretched over the entire external surface of the bone, including both the superstructures and the growth plates, it can pass to the growth plates signals concerning the relative position of superstructures.”<-Then perhaps we can manipulate longitudinal bone growth by manipulating the periosteal sheath.
“periosteal tension down-regulates growth plate activity, as the higher the tension level, the more inhibited growth plate activity is. Damaged periosteum forms a scar tissue at the site of destruction. This scar tissue, which anchors the periosteum into the bone, creates an independent tension level near each growth plate. As a result, a new growth balance is formed, which equals the ratio between the distances from the site of the scar to the two ends of the bone, therefore maintaining the relative position of the scar site.. Superstructures can be considered as natural anchoring points for the periosteum into the ossified bone, either due to the insertion of tendons through them into the bone cortex, or by means of steric interference, such as in the tibiofibular junction. This results in a regulatory loop whereby the superstructures determine the tension levels of the two periosteal segments, which control the ratio of growth rates by inhibiting growth plate activity, which in turn maintains the relative position of the superstructure.”
Mechanical regulation of musculoskeletal system development
“During embryogenesis, the musculoskeletal system develops while containing within itself a force generator in the form of the musculature. This generator becomes functional relatively early in development, exerting an increasing mechanical load on neighboring tissues as development proceeds. A growing body of evidence indicates that such mechanical forces can be translated into signals that combine with the genetic program of organogenesis. This unique situation presents both a major challenge and an opportunity to the other tissues of the musculoskeletal system, namely bones, joints, tendons, ligaments and the tissues connecting them. Here, we summarize the involvement of muscle-induced mechanical forces in the development of various vertebrate musculoskeletal components and their integration into one functional unit.”
“These forces [on the cell cytoskeleton], which can be translated into biochemical signals by molecules possessing mechanotransduction capabilities, are transmitted across transmembrane receptors into the extracellular matrix (ECM) and can also reach neighboring cells.”
“In the case of musculoskeletal development, exogenous forces acting on tendons and the skeleton are generated by muscle contraction.”
“Bone morphology is regulated by mechanical forces at different levels, as demonstrated by the various developmental and functional aberrations that arise in the absence of muscle contraction. (1) Bone elongation is impaired due to reduced chondrocyte proliferation in the growth plate. (2) Additionally, the organization of resting chondrocytes into columns is impaired, which can also affect skeletal elongation. (3) Bone eminence growth is arrested, resulting in smaller or absent eminences. (4) Differential appositional growth is lost, resulting in a circular circumferential shape. (5) Joint formation is impaired during embryonic development, leading to joint fusion.”
“These effects on chondrocyte proliferation could be mediated by yes-associated protein 1 (YAP1), a mechanosensor that is part of the Hippo signaling pathway. Indeed, changes in YAP cellular localization in chondrocytes were identified in vitro in response to matrix stiffness, and YAP was shown to regulate bone size, promote chondrocyte proliferation and inhibit chondrocyte differentiation in vitro and in vivo by suppressing collagen type X”
“mechanical forces can regulate both the content and the dynamics of proteoglycan and collagen production by these cells”
“Bones grow in width by preferential periosteal growth, which involves repetitive steps of strut-and-ring construction by mineral deposition.”
“non-selective mechanosensitive cationic channels, PIEZO1 and PIEZO2, which are expressed in a variety of tissue types, including chondrocytes”
“the attachment between the very elastic tendon and the very rigid bone creates a point of high stress concentration during force transfer, which could lead to detachment. Dissipation of this stress is achieved either by the formation of fibrous attachments, in which tendon fibers are inserted into the cortical bone in a structure that resembles a root system, or by the formation of a fibrocartilaginous attachment composed of different layers that gradually change in stiffness. Although enthesis development begins in the embryo, the formation of the unique transitional tissue and its subsequent mineralization occur postnatally.”
“In the mature skeleton, the tendon-bone enthesis is an interfacial zone of transitional tissue located between compliant, fibrous tendon to rigid, dense mineralized bone. This transitional tissue provides a mechanism of stress and strain reduction at the interface between two mechanically dissimilar tissues”
“Fibrous entheses are generally found at insertion sites of stabilizing tendons, whereas fibrocartilaginous entheses are typically found at insertions of tendons that contribute to joint movement. Fibrous enthesis attach directly to bone and typically form Sharpey’s fibers, which are perforating fibers that embed into bone’s periosteal surface”
“Fibrocartilaginous entheses consist of four distinct histological zones, including aligned tendon, unmineralized fibrocartilage, mineralized fibrocartilage, and subchondral bone. A smooth and uniform basophilic tidemark distinguishes the transition between the two fibrocartilaginous zones, and this tidemark is disrupted and irregular in enthesopathy. The fibrocartilage enthesis matures during postnatal growth in response to mechanical loads from skeletal muscle and consists of cells that express both tenogenic and chondrogenic factors”
“Morphologically, the development of the enthesis has been likened to a “miniature” or arrested growth plate. However, unlike growth plates in long bones which eventually fuse at skeletal maturity, the fibrocartilage of the enthesis retains the morphological features of fibrocartilage and maintain Gli1+ cells at the interface throughout postnatal growth”
Lasting organ-level bone mechanoadaptation is unrelated to local strain
“Bones adapt to mechanical forces according to strict principles predicting straight shape. Most bones are, however, paradoxically curved. To solve this paradox, we used computed tomography–based, four-dimensional imaging methods and computational analysis to monitor acute and chronic whole-bone shape adaptation and remodeling in vivo. We first confirmed that some acute load-induced structural changes are reversible, adhere to the linear strain magnitude regulation of remodeling activities, and are restricted to bone regions in which marked antiresorptive actions are evident.{this is actually a good finding because it emphasizes that bone is an adaptive tissue} We make the novel observation that loading exerts significant lasting modifications in tibial shape and mass across extensive bone regions, underpinned by (re)modeling independent of local strain magnitude, occurring at sites where the initial response to load is principally osteogenic. This is the first report to demonstrate that bone loading stimulates nonlinear remodeling responses to strain that culminate in greater curvature adjusted for load predictability without sacrificing strength.”
“Wolff’s law is at odds with the curved overall shape of most bones, however, because adherence to these principles would predict straighter bones.”
“bones is instead optimized for load predictability. This would lead to the prediction that as bone curvature increases and predictability of the bending direction is augmented, its strength decreases”
“bone acts as an organ to acquire lasting modifications in shape and strength with greater bending predictability, which involves coordination of spatial remodeling that is unrelated to local strain magnitude.”<-it could be unrelated to local strain magnitude because of fluid forces.
“acute bone gains are lost within 6 weeks after load and extend this by showing that this is achieved through a simple reversal, involving greater resorption and lower formation in the proximal tibia in the chronic phase”
“net load-induced acute gains in the mid-to-distal tibia are, in contrast, lasting. Our 4D analyses reveal that this gain in bone mass is achieved predominantly by an enhanced formation in the acute phase, with only minor subsequent modifications in remodeling in the chronic postload phase. Comparing the remodeling activities in these two diverse, proximal and distal, locations by our novel application of whole-bone 4D analyses leads us to propose that the rapidity of load-induced acute changes in bone remodeling and the rate of their chronic postload reversal are linked. Reversibility in the proximal tibia is aligned to a marked load-related antiresorptive effect, while, contrarily, the lasting adaptation in the mid-to-distal region is principally linked with load-induced osteogenesis. It is intriguing that only the latter leads to permanent modification in bone shape.”
“Long-term curvature changes in nonbiological materials can involve creep deformation.”
“We show that some acute load-induced structural changes are reversible and adhere to linear strain magnitude feedback-loop regulation of remodeling activities. On the other hand, a vast proportion of the bone retains lasting structural memory of loading to generate nonlinear, strain magnitude–independent remodeling to achieve greater curvature optimized for load predictability without sacrificing strength.”<-some bone changes may be due to creep deformation thus being permanent.
Heterotopic ossification is endochondral ossification that occurs outside the bone. Understanding why it occurs can help us find ways to induce endochondral ossification within the bone. The biggest issue with inducing a new growth plate in bone is the permissive local environment criteria. The bone likely has to be degraded in some way to induce a neo-growth plate as the existing bone environment likely puts a constraining factor on growth.
“Heterotopic ossification (HO) is a debilitating condition defined by the de novo development of bone within non-osseous soft tissues, and can be either hereditary or acquired. The hereditary condition, fibrodysplasia ossificans progressiva is rare but life threatening. Acquired HO is more common and results from a severe trauma that produces an environment conducive for the formation of ectopic endochondral bone. Despite continued efforts to identify the cellular and molecular events that lead to HO, the mechanisms of pathogenesis remain elusive. It has been proposed that the formation of ectopic bone requires an osteochondrogenic cell type, the presence of inductive agent(s) and a permissive local environment. To date several lineage-tracing studies have identified potential contributory populations. However, difficulties identifying cells in vivo based on the limitations of phenotypic markers, along with the absence of established in vitro HO models have made the results difficult to interpret. The purpose of this review is to critically evaluate current literature within the field in an attempt identify the cellular mechanisms required for ectopic bone formation. The major aim is to collate all current data on cell populations that have been shown to possess an osteochondrogenic potential and identify environmental conditions that may contribute to a permissive local environment. This review outlines the pathology of endochondral ossification, which is important for the development of potential HO therapies and to further our understanding of the mechanisms governing bone formation.”
“of the 80 % of war victims who suffer major extremity trauma during combat injury, approximately 64 % of these patients go on to develop some degree of HO”
“Current evidence suggests that the formation of ectopic bone in vivo requires three primary conditions: (1) a cell type capable of osteogenic differentiation, (2) the presence of inductive agents and (3) a permissive local environment”
“o date many contributory biological factors have been implicated in the aetiology, including the bone morphogenetic proteins (BMPs), inflammation, prostaglandin E2, hypercalcemia, hypoxia, abnormal nerve activity, immobilisation and dysregulation of hormones”
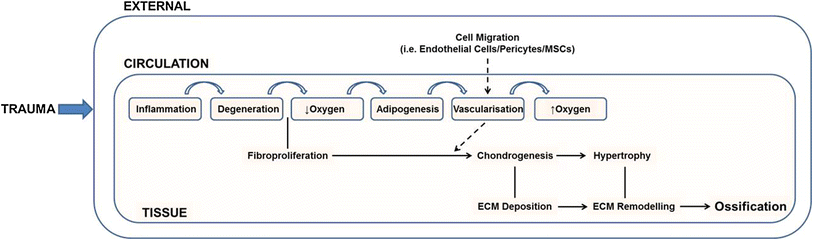
“Tissue damage leads to the infiltration of immunological cells (monocytes, neutrophils and leukocytes) through the local vasculature. Resulting fibro-proliferation of an as yet unknown cell population is accompanied by hypoxia and the generation of brown adipose tissue at the site of damage. The presence of adipose tissue is hypothesised to lower the local oxygen tension leading to the establishment of a chondrogenic environment. Neovascularisation accompanies chondrogenesis and provides an avenue through which systemic cell types (endothelial cells, pericytes etc.,) may enter the injury site, and potentially contributed to osteochondrogenic differentiation. A subsequent increase in local oxygen tension promotes chondrocyte maturation and hypertrophy. The collagenous matrix deposited by these cells is then remodelled and ossified to form endochondral bone”<-If we induce such factors in the bone we can create new growth plates in there too.
“MSCs have frequently been shown to form endochondral bone when cultured under appropriate conditions (e.g. under hypoxia and/or in the presence of TGF-β)”
“MSCs may also contribute to chondrocyte hypertrophy and the progression of HO via their immunomodulatory effects, primarily through the production of anti-inflammatory cytokines and nitric oxide (NO)”
Several cell types are listed that are capable of heterotopic ossification are likely present in bone.
“Bone marrow HSC side population Lin−/Sca-1+/cKit+/CD45+”
“cells presenting the glutamate transporter GLAST were found to contribute to the formation of ectopic bone, and that these GLAST+ cells appeared to be distinct from the Tie2+ population”
” a significant upregulation in transcriptional activity in key osteogenesis-related genes (ALPL, BMP-2, BMP-3, COL2A1, COLL10A1, COL11A1, COMP, CSF2, CSF3, MMP8, MMP9, SMAD1 and VEGFA) in patients that developed HO compared to those who did not.”
“Heterotopic ossification (HO) is a metaplastic biological process in which there is newly formed bone in soft tissues, resulting in joint mobility deficit and pain. Different treatment modalities have been tried to prevent HO development, but there is no consensus on a therapeutic approach. Since electrical stimulation is a widely used resource in physiotherapy practice to stimulate joint mobility, with analgesic and anti-inflammatory effects, its usefulness for HO treatment was investigated. We aimed to identify the influence of electrical stimulation on induced HO in Wistar rats. Thirty-six male rats (350-390 g) were used, and all animals were anesthetized for blood sampling before HO induction, to quantify the serum alkaline phosphatase. HO induction was performed by bone marrow implantation in both quadriceps of the animals, which were then divided into 3 groups: control (CG), transcutaneous electrical nerve stimulation (TENS) group (TG), and functional electrical stimulation (FES) group (FG) with 12 rats each. All animals were anesthetized and electrically stimulated twice per week, for 35 days from induction day. After this period, another blood sample was collected and quadriceps muscles were bilaterally removed for histological and calcium analysis and the rats were killed. Calcium levels in muscles showed significantly lower results when comparing TG and FG (P<0.001) and between TG and CG (P<0.001). Qualitative histological analyses confirmed 100% HO in FG and CG, while in TG the HO was detected in 54.5% of the animals. The effects of the muscle contractions caused by FES increased HO, while anti-inflammatory effects of TENS reduced HO.”
“The formation of heterotopic bone may be due to muscle trauma (myositis ossificans). It is common in people who have undergone total hip arthroplasty , those with spinal cord injuries, and victims of head trauma, all of which often lead to long periods of immobilization of the affected limbs.”
“skeletal muscle serves as a physical safeguard for the other organs and is anatomically located immediately beneath the skin, so it represents the most damaged organ in the body. Although skeletal muscle is characterized by the presence of fatty and connective tissues that originated from nonmyogenic mesenchymal progenitors, those progenitors were initially identified in BM”
“Muscle contraction occurs by the deposition of calcium in muscle tissue, and this stimulates the sliding of actin and myosin myofibrils, which characterizes the contractile process”
“electrical stimulation helps the deposition of calcium, causes changes in oxygen content and pH, stimulates expression of growth factors, and recruits help in osteoblast migration and secretion of extracellular matrix (ECM), leading to bone formation.”
“Mechanotransduction refers to the process by which the body converts a mechanical stimulus into a cellular response”
“Osteoarthritis (OA) is associated with the metabolic syndrome, however the underlying mechanisms remain unclear. We investigated whether low density lipoprotein (LDL) accumulation leads to increased LDL uptake by synovial macrophages and affects synovial activation, cartilage destruction and enthesophyte/osteophyte formation during experimental OA in mice.
LDL receptor deficient (LDLr−/−) mice and wild type (WT) controls received a cholesterol-rich or control diet for 120 days. Experimental OA was induced by intra-articular injection of collagenase twelve weeks after start of the diet. OA knee joints and synovial wash-outs were analyzed for OA-related changes. Murine bone marrow derived macrophages were stimulated with oxidized LDL (oxLDL), whereupon growth factor presence and gene expression were analyzed.
A cholesterol-rich diet increased apolipoprotein B (ApoB) accumulation in synovial macrophages. Although increased LDL levels did not enhance thickening of the synovial lining, S100A8 expression within macrophages was increased in WT mice after receiving a cholesterol-rich diet, reflecting an elevated activation status. Both a cholesterol-rich diet and LDLr deficiency had no effect on cartilage damage; in contrast, ectopic bone formation was increased within joint ligaments (fold increase 6.7 and 6.1, respectively). Moreover, increased osteophyte size was found at the margins of the tibial plateau (4.4 fold increase after a cholesterol-rich diet and 5.3 fold increase in LDLr−/− mice). Synovial wash-outs of LDLr−/− mice and supernatants of macrophages stimulated with oxLDL led to increased transforming growth factor-beta (TGF-β) signaling compared to controls.
LDL accumulation within synovial lining cells leads to increased activation of synovium and osteophyte formation in experimental OA. OxLDL uptake by macrophages activates growth factors of the TGF-superfamily.”
“multiple injections of members of the TGF-super family, such as TGF-β or BMP-2, directly into the knee joint of the mouse caused abundant enthesophyte/osteophyte formation”

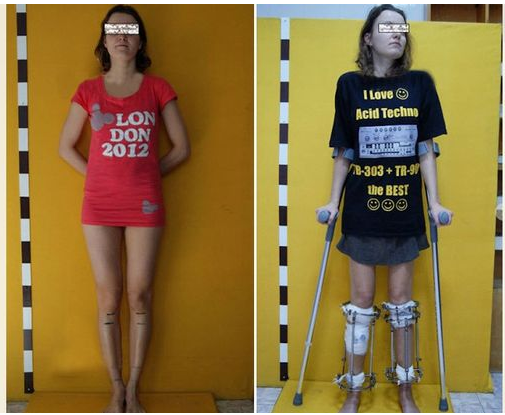 This is sort of old news but it is sometimes still worth mentioning. Back in March of this year there was another story that came out of a person who got cosmetic limb lengthening surgery. Unlike most of the people you hear about, this person was a girl.
This is sort of old news but it is sometimes still worth mentioning. Back in March of this year there was another story that came out of a person who got cosmetic limb lengthening surgery. Unlike most of the people you hear about, this person was a girl.