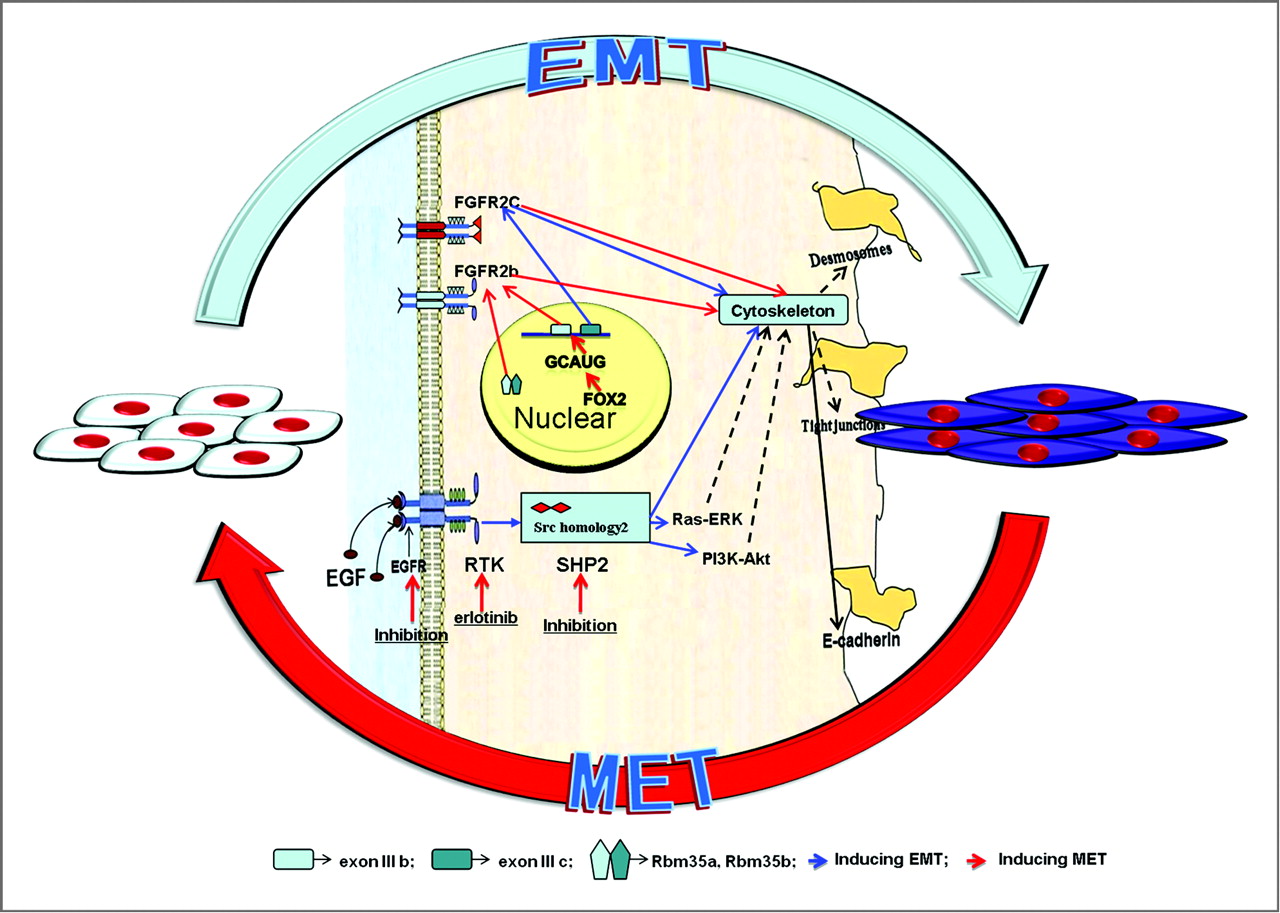
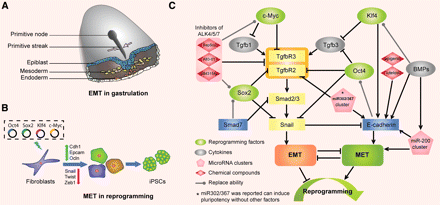
Here’s a review paper on factors that alter the bone microenvironment. There’s a lot of stuff here. More than is mentioned.
Biomechanical and biophysical environment of bone from the macroscopic to the pericellular and molecular level.
” Bones with complicated hierarchical configuration and microstructures constitute the load-bearing system. Mechanical loading plays an essential role in maintaining bone health and regulating bone mechanical adaptation (modeling and remodeling). The whole-bone or sub-region (macroscopic) mechanical signals, including locomotion-induced loading and external actuator-generated vibration, ultrasound, oscillatory skeletal muscle stimulation, etc., give rise to sophisticated and distinct biomechanical and biophysical environments at the pericellular (microscopic) and collagen/mineral molecular (nanoscopic) levels, which are the direct stimulations that positively influence bone adaptation. While under microgravity, the stimulations decrease or even disappear, which exerts a negative influence on bone adaptation. A full understanding of the biomechanical and biophysical environment at different levels is necessary for exploring bone biomechanical properties and mechanical adaptation. In this review, the mechanical transferring theories from the macroscopic to the microscopic and nanoscopic levels are elucidated. First, detailed information of the hierarchical structures and biochemical composition of bone, which are the foundations for mechanical signal propagation, are presented. Second, the deformation feature of load-bearing bone during locomotion is clarified as a combination of bending and torsion rather than simplex bending. The bone matrix strains at microscopic and nanoscopic levels directly induced by bone deformation are critically discussed, and the strain concentration mechanism due to the complicated microstructures is highlighted. Third, the biomechanical and biophysical environments at microscopic and nanoscopic levels positively generated during bone matrix deformation or by dynamic mechanical loadings induced by external actuators, as well as those negatively affected under microgravity, are systematically discussed, including the interstitial fluid flow (IFF) within the lacunar-canalicular system and at the endosteum, the piezoelectricity at the deformed bone surface, and the streaming potential accompanying the IFF. Their generation mechanisms and the regulation effect on bone adaptation are presented. The IFF-induced chemotransport effect, shear stress, and fluid drag on the pericellular matrix are meaningful and noteworthy. Furthermore, we firmly believe that bone adaptation is regulated by the combination of bone biomechanical and biophysical environment, not only the commonly considered matrix strain, fluid shear stress, and hydrostatic pressure, but also the piezoelectricity and streaming potential. Especially, it is necessary to incorporate bone matrix piezoelectricity and streaming potential to explain how osteoblasts (bone formation cells) and osteoclasts (bone resorption cells) can differentiate among different types of loads. Specifically, the regulation effects and the related mechanisms of the biomechanical and biophysical environments on bone need further exploration, and the incorporation of experimental research with theoretical simulations is essential.”
“Osteocyte processes are attached to the canalicular wall via transverse tethering elements
(the average spacing between the elements is 32 ± 14 nm) and only directly contact the canalicular wall at specialized collagen fibril protrusions ”
“In trabecular bone, the trabecular rods and plates form three-dimensional structures, and within the trabeculae are less regular arranged lamellae and osteocytes”
“The organic material, which is primarily type I collagen (90%) and nonfibrillar organic matrix
(osteocalcin and osteopontin present in a large proportion) gives the bone toughness. The mineral salts, mainly nanocrystallite apatite materials, permeate the organic matrix and provide the characteristic rigidity and strength of bone. Common belief holds that the apatite materials are hydroxyapatite [Ca10(PO4)6(OH)2] (HAP). In fact, there is no OH-band in bone mineral detected in a Raman spectrum, therefore the bone apatite is not hydroxyapatite ”
“in two adults tibiae during comfortable walking and strenuous running; the results suggested that the compressive strain ranged from approximately 400 to 1300 µε, and the tensile strain ranged from approximately 380 to 750 µε ”
“The pericellular space in the lacunar-canalicular system (LCS) of cortical bone and the porous
structures within trabecular bone are filled with interstitial fluid. Due to the low permeability of mineralized bone matrix, interstitial fluid flow (IFF) is principally generated during alteration of intramedullary pressurization (ImP) and bone matrix deformation”
“Pulsatile extravascular pressure resulting from blood circulation can also drive IFF, but it is much smaller than that induced by ImP and bone deformation “<- so maybe muscular contraction is not as important as thought and that’s why bodybuilders have not been shown to grow taller as a result of bodybuilding.
“The IFF generated from ImP usually follows two spatially distinct flow profiles: IFF within the
LCS and at the endosteal surface. Uniform pressurization [such as that due to decreased intramedullary cavity resulting from elevated bone marrow lipids induced by high level of corticosteroid administration will generate radial flow from the intramedullary compartment to the endosteal surface and into the LCS due to the pressure gradient from the marrow cavity to the bone matrix. Non-uniform pressure gradients within the intramedullary cavity [such as those due to local heterogeneous permeability or fluid displacement changes in the intramedullary compartment from the interaction between mechanical loading/oscillatory muscle stimulation and capillary filtration in bone tissue will cause tangential fluid flow to the endosteal surface ”
“Because the mineralized matrix is nearly impermeable{This is partially why it is so hard to grow taller} and the pericellular annular fluid space surrounding the osteocyte cell body and processes is narrow (gap < 1 µm) , the transportation of nutrients (e.g., glucose), oxygen, waste products (e.g., lactic acid), and regulatory signals (e.g., sex hormones, nitric oxide, prostaglandins, cytokines, and growth factors) by simple diffusion is not sufficient to maintain cell viability and function IFF within the LCS serves as the primary transport mechanism between the blood supply and osteocytes”
“hydrostatic pressure is induced by the bone compression that results from physiological loading. Based on the data that walking at a frequency of 1 Hz induces bone strain of 1000 µε and 0–18 MPa cyclic stress{this is surprisingly high}, the maximum hydrostatic pressure in LCS is approximately 0.27 MPa{this is actually within the chondrogenic range of 0.1-10MPa; there must be additional factors going on}, this pressure is estimated to be 12% of the applied axial stress and 40 times higher than that in the vasculature”
” the compressed areas develop negative potentials, whereas the tensioned areas develop positive potentials. These changes are known as the direct piezoelectric effect ”
“The researchers subjected demineralized type I collagen to bending deformation and bathed the collagen in continually flowing physiologic fluid over a period of weeks. Interestingly, the compressed internal surface (negative) of the collagen attracted calcium ions (positive), leading to the subsequent nucleation and crystallization of apatite and an increase in precipitate. In contrast, the tensioned portion (positive) of the collagen showed no difference in precipitate compared with that in the undeformed collagen. ”
“Apatite is piezoelectric by nature ”
“dry collagen is piezoelectric, but fully hydrated collagen is not piezoelectric because the structured water may increase the symmetry of the collagen molecule”
” In the presence of apatite, access of water to parts of the collagen fibril and the collagen swelling to its fully hydrated form can be prevented, thus the collagen can show some piezoelectricity even when the bone is fully hydrated ”
” the negative and positive potentials caused by polarization can be stored in apatite for years ”
“osteons are irregular cylinders always with complex branching and interconnections”
“The strain-induced movement of interstitial fluid through bone porosities can induce another
bioelectrical potential in addition to piezoelectricity; this potential is called the streaming potential ”
“At the interface of the bone matrix and interstitial fluid, negative charges accumulate on the bone surface due to the electronegativity of collagen; these charges attract positive ions in the liquid, and the positive ions concentrate near the interface, forming an electrical double layer. The electric potential in the interfacial double layer relative to the interstitial fluid away from the interface is the zeta potential (ζ), and when interstitial fluid flows tangentially along the interface under pressure, a streaming potential appears along the flow direction. The zeta potential is mainly generated by deformed collagen and corresponds to the streaming potential ”
” the body fluid shifts[during space flight], and perfusion is diminished in the lower limbs and increased in the upper limbs and head ”
“ultrasound may primarily exert an effect on increasing blood flow and producing micromechanical strains; the actual acoustic intensity experienced by the bone cells at the target site might be lower due to the possible attenuation caused by the overlying soft tissue ”
“[Oscillatory muscle stimulation] at 20 Hz generated a maximum ImP and a relative high bone strain”