
One of the key parts of LSJL is to cause new growth plate formation via inducing differentiation of epiphyseal bone marrow. This study has insight that epiphyseal bone marrow may participate in the growth plate process and thus in turn may be capable of contributing to neo-growth plate formation.
“Because of its close relationship to tetrapods{A tetrapod is a four limbed organism}, Eusthenopteron{a kind of fish} is an important taxon for understanding the establishment of the tetrapod body plan. Notably, it is one of the earliest sarcopterygians in which the humerus of the pectoral fin skeleton is preserved. The microanatomical and histological organization of this humerus provides important data for understanding the evolutionary steps that built up the distinctive architecture of tetrapod limb bones. Previous histological studies showed that Eusthenopteron’s long-bone organization was established through typical tetrapod ossification modalities. Based on a three-dimensional reconstruction of the inner microstructure of Eusthenopteron’s humerus, obtained from propagation phase-contrast X-ray synchrotron microtomography, we are now able to show that, despite ossification mechanisms and growth patterns similar to those of tetrapods, it also retains plesiomorphic{trait that only eusthenopteron’s have but not other tetrapods} characters such as a large medullary cavity, partly resulting from the perichondral ossification around a large cartilaginous bud as in actinopterygians. It also exhibits a distinctive tubular organization of bone-marrow processes. The connection between these processes and epiphyseal structures highlights their close functional relationship, suggesting that either bone marrow played a crucial role in the long-bone elongation processes or that trabecular bone resulting from the erosion of hypertrophied cartilage created a microenvironment for haematopoietic stem cell niches. ”
Now even though these fish and humans are different it suggests still that bone marrow has a capability to form new growth plates.
Here’s the anatomy of the mid-bone shaft of this fish(juvenile):
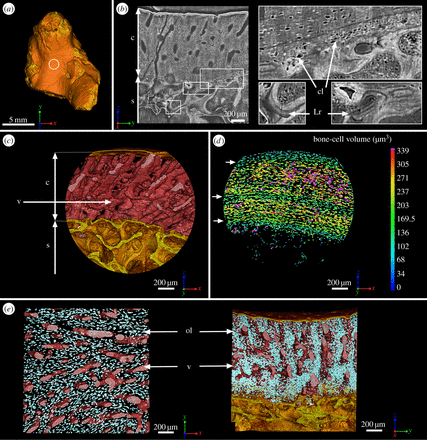
“(b) Virtual thin section (made along the longitudinal axis) showing the primary bone deposit of cortical bone and its connection to the spongiosa. Some remnants of calcified cartilage are still preserved at the location of Katschenko’s line (chondrocyte lacunae) and within the spongiosa (Liesegang rings).”
So “micro-growth plates” within the bone marrow.
“At mid-shaft (figure 1a), the inner surface of the cortical bone is delimited by clusters of numerous large globular cell lacunae (cl, figure 1b,d) that can be identified as chondrocyte lacunae of cartilage. This suggests that remnants of Katschenko’s line are still present. Several stacks of Liesegang rings, typical of calcified cartilage, are also notable among the endochondral trabeculae”
“In extant tetrapods, the spongiosa forms when chondroclasts create erosion bays in the cartilage that are then lined with a thin peripheral bone layer, and it is common for small remnants of calcified cartilage to be left behind by the process; spongiosa formation in Eusthenopteron appears to have been similar.”
“The trabeculae in the spongiosa [of adult fish] are greatly remodelled and show no visible remnant of calcified cartilage”
“In extant actinopterygians, the fin endoskeleton develops from a blastema that differentiates into cartilages. Perichondral bone is deposited on the surface of the cartilages and continues to grow centrifugally as periosteal bone”
“a typical metapterygial bone is composed of a cartilaginous rod surrounded by a bony tube with cartilage projecting as condyles”
“In contrast to tetrapods, actinopterygian long bones have no haematopoietic bone marrow but only fatty tissues in the spaces created by the erosion of the cartilaginous rod. Neither do they show any evidence of a growth plate with longitudinally oriented columns of chondrocytes. The large longitudinal tubular mesh observed in Eusthenopteron humerus appears to constitute the earliest and phylogenetically deepest documented occurrence of a complex functional bone marrow in the tetrapod stem group.”
“As the tubular channels in Eusthenopteron are obviously connected to the epiphyses, the appearance of a complex bone marrow seems to be related to the appearance of tetrapod-like epiphyseal structures and elongation growth. Eusthenopteron lacks the comprehensive remodelling and trabecular resorption that creates an open medullary cavity in the majority of extant tetrapods, but the reduction of trabeculae between the longitudinal tubes in the adult compared with the juvenile may represent an evolutionary precursor of this process. ”
“Tetrapod bone marrow has been shown capable of degrading cartilage proteoglycans and inducing the initial stage of endochondral ossification”