
Knowledge about the attachment of ligaments and tendons to bone is important to the new method of LSJL loading. This method involves clamping regions where two bones are attached to each other via ligaments.
According to Physical Therapies in Sport and Exercise there is no communication between tendons and bone however. It describes the tendon enthesis as as an unmineralized bundle of rounded cell fibrocartilage wrapped in bundles of type II collagen. It states though that these cells do not have connective arms to other cells so tendons and bones would have no communication. We are interested in ligaments rather than tendons, as tendons would get a lot of load from muscular activity but is still relevant as both tendons and ligaments have entheses.
Enthesis fibrocartilage cells originate from a population of Hedgehog-responsive cells modulated by the loading environment.
“Tendon attaches to bone across a specialized tissue called the enthesis. This tissue modulates the transfer of muscle forces between two materials, i.e. tendon and bone, with vastly different mechanical properties. The enthesis for many tendons consists of a mineralized graded fibrocartilage that develops postnatally, concurrent with epiphyseal mineralization. By genetically demarcating cells expressing Gli1 in response to Hedgehog (Hh) signaling, we discovered a unique population of Hh-responsive cells in the developing murine enthesis that were distinct from tendon fibroblasts and epiphyseal chondrocytes. Lineage-tracing experiments revealed that the Gli1 lineage cells that originate in utero eventually populate the entire mature enthesis. Muscle paralysis increased the number of Hh-responsive cells in the enthesis, demonstrating that responsiveness to Hh is modulated in part by muscle loading{surprisingly lower muscle loading seems to increase responsiveness}. Ablation of the Hh-responsive cells during the first week of postnatal development resulted in a loss of mineralized fibrocartilage, with very little tissue remodeling 5 weeks after cell ablation. Conditional deletion of smoothened, a molecule necessary for responsiveness to Ihh, from the developing tendon and enthesis altered the differentiation of enthesis progenitor cells, resulting in significantly reduced fibrocartilage mineralization and decreased biomechanical function. Hh signaling within developing enthesis fibrocartilage cells is required for enthesis formation.”
“Fibrous entheses include a periosteal component and typically insert into the metaphyses or diaphyses of long bones. By contrast, fibrocartilagenous attachments typically insert into epiphyses or bone ridges. These attachments are characterized by a partially mineralized region of fibrocartilage at the tendon-bone interface, with a round cell morphology and increased proteoglycan content relative to tendon. Due to these characteristics, mature entheses have been likened to arrested growth plates“
Since tendons at first glance do not have much communication between themselves and bones how about ligaments?
Establishing a Coculture System for Ligament-Bone Interface Tissue Engineering
“Ligament-bone interface (enthesis) is a complex structure which comprises of ligament, fibrocartilage and bone. The fibrocartilage transformation adds significant insertional strength to the interface and makes it highly resistant to avulsion forces. Many ACL grafts cannot generate native interfacial region, leading to their failure. Co-culture has proved to be an effective way to generate new tissues in tissue engineering. Studies have found important signaling molecules in transduction pathway of chondrogenesis to be transmitted via gap junctions. Stem cells cocultured between ligament and bone cells would enable transmission of chondrogenic factors from bone/ligament cells to bone marrow stem cells (BMSCs) via gap junctions, resulting in their differentiation into fibrocartilage. In this study, two set of co-culture (BMSCs and ligament cells; BMSCs and bone cells) were established. Confocal microscopy showed efficient dye transfer from bone/ligament cells into BMSCs. This was further confirmed and quantified by FACS, which showed a gradual temporal increase in the percentage of BMSCs acquiring Calcein.”
This study also states that there is ligament and bone communication:
In vitro study of stem cell communication via gap junctions for fibrocartilage regeneration at entheses.
“Entheses are fibrocartilaginous organs that bridge ligament with bone at their interface and add significant insertional strength. To replace a severely damaged ligament, a tissue-engineered graft preinstalled with interfacial fibrocartilage, which is being regenerated from stem cells, appears to be more promising than ligament-alone graft. Such a concept can be realized by a biomimetic approach of establishing a dynamic communication of stem cells with bone cells and/or ligament fibroblasts in vitro.
The current study has two objectives. The first objective is to demonstrate functional coculture of bone marrow-derived stem cells (BMSCs) with mature bone cells/ligament fibroblasts as evidenced by gap-junctional communication in vitro. The second objective is to investigate the role of BMSCs in the regeneration of fibrocartilage within the coculture.
Rabbit bone/ligament fibroblasts were dual-stained with DiI-Red and calcein (gap-junction permeable dye), and cocultured with unlabeled BMSCs at fixed ratio (1:10). The functional gap junction was demonstrated by the transfer of calcein from donor to recipient cells that was confirmed and quantified by flow cytometry. Type 2 collagen (cartilage extracellular matrix-specific protein) expressed by the mixed cell lines in the cocultures were estimated and compared with that of the ligament-bone coculture (control).
Significant transfer of calcein into BMSCs was observed and flow cytometry analyses showed a gradual increase in the percentage of BMSCs acquiring calcein with time. Cocultures that included BMSCs expressed significantly more type 2 collagen compared with the control.
The current study reported the expression of gap-junctional communication of BMSCs with two adherent cell lines of musculoskeletal system in vitro and also confirmed that incorporation of stem cells augments fibrocartilage regeneration.”
“Inside the embryo, tendons and ligaments appear as fibrous cellular outgrowths from the perichondrium of cartilaginous precursor to later bone. In the neonates, as the cartilaginous anlage ossifies, fibrocartilaginous elements of the ligament are cemented into the initial calcified cartilage, which is later replaced by mature bone. Only a thin layer of proliferative epiphyseal cartilage remains at the interface{this is a very promising fact as proliferative epiphyseal cartilage is a solid start in term of neo growth plate formation}. The resulting mature tendon or ligament insertion comprises four zones: collagenous ligament or tendon, nonmineralized fibrocartilage, mineralized fibrocartilage and cancellous bone”
The role of muscle loading on bone (Re)modeling at the developing enthesis.
“Muscle forces are necessary for the development and maintenance of a mineralized skeleton. Removal of loads leads to malformed bones and impaired musculoskeletal function due to changes in bone (re)modeling. In the current study, the development of a mineralized junction at the interface between muscle and bone was examined under normal and impaired loading conditions. Unilateral mouse rotator cuff muscles were paralyzed using botulinum toxin A at birth. Control groups consisted of contralateral shoulders injected with saline and a separate group of normal mice. It was hypothesized that muscle unloading would suppress bone formation and enhance bone resorption at the enthesis, and that the unloading-induced bony defects could be rescued by suppressing osteoclast activity. In order to modulate osteoclast activity, mice were injected with the bisphosphonate alendronate. Bone formation was measured at the tendon enthesis using alizarin and calcein fluorescent labeling of bone surfaces followed by quantitative histomorphometry of histologic sections. Bone volume and architecture was measured using micro computed tomography. Osteoclast surface was determined via quantitative histomorphometry of tartrate resistant acid phosphatase stained histologic sections. Muscle unloading resulted in delayed initiation of endochondral ossification at the enthesis, but did not impair bone formation rate. Unloading led to severe defects in bone volume and trabecular bone architecture. These defects were partially rescued by suppression of osteoclast activity through alendronate treatment, and the effect of alendronate was dose dependent. Similarly, bone formation rate was increased with increasing alendronate dose across loading groups. The bony defects caused by unloading were therefore likely due to maintained high osteoclast activity, which normally decreases from neonatal through mature timepoints. These results have important implications for the treatment of muscle unloading conditions such as neonatal brachial plexus palsy, which results in shoulder paralysis at birth and subsequent defects in the rotator cuff enthesis and humeral head. “
This study only studied tendon and not ligament entheses though.
This site has a lot of into about entheses:
“The normal enthesis has a tendency to develop new bone formation with ageing. This manifests as incidental “bony spurs” that are visible on X-rays”
“Joint tissues under tension favours bone development and the outermost part of the enthesis is subject to predominant tension (pulling force).”
Here’s an entheses with neo-bone formation: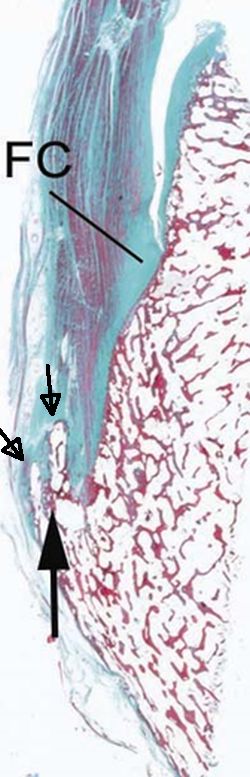 Fc stands for fibrocartilage and the arrows are pointing at abnormal bone formation. Not how the fibrocartilage is shaped into the bone like a zone of ranvier.
Fc stands for fibrocartilage and the arrows are pointing at abnormal bone formation. Not how the fibrocartilage is shaped into the bone like a zone of ranvier.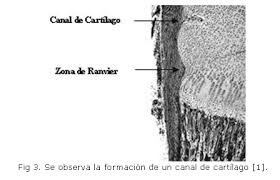
This website describes the groove of Ranvier:
“During the first year of life, the zone spreads over the adjacent metaphysis to form a fibrous circumferential ring bridging from the epiphysis to the diaphysis”
“This ring increases the mechanical strength of the physis and is responsible for appositional bone growths. Supplies chondrocytes to periphery“
The site also says that ligaments have rounder cell morphology than ligaments which is more consistent with a chondrocyte cell. Interestingly it says that ligaments also do not plastically deform which means that they can not permanently increase in length as a result of pulling force.
Strategies to engineer tendon/ligament-to-bone interface: Biomaterials, cells and growth factors.
“The complex and heterogeneous structure of the enthesis is essential to ensure smooth mechanical stress transfer between bone and soft tissues. Following injury, the interface is not regenerated, resulting in high rupture recurrence rates. “
“entheses exhibit gradients in tissue organization, composition and mechanical properties that serve to (1) effectively transfer stress between mechanically dissimilar materials and (2) sustain the heterotypic cellular communications required for interface function and homeostasis”
“the first part of the enthesis consists of fibrous connective tissue (tendon/ligament) characterized by the presence of parallel collagen fibers with interspaced elongated fibroblasts organized in arrays. The ECM primarily consists of collagen type I and small amounts of proteoglycans. The next zone is non-mineralized fibrocartilage, populated by round fibrochondrocytes arranged in rows. The ECM contains mostly collagen type II (typically characteristic of hyaline cartilage) as well as high levels of pericellular collagen type III, small amounts of collagen types I and X, and proteoglycans (mainly aggrecan) with associated chondroitin 4- and 6-sulfate glycosaminoglycans (GAGs). The third zone is mineralized fibrocartilage. This zone is populated by hypertrophic fibrochondrocytes, which are round and bigger than fibrochondrocytes. The ECM is composed mainly of collagen type II with significant amounts of collagen type X and aggrecan. Finally, the mineralized fibrocartilage merges into bone tissue containing osteoblasts, osteocytes and osteoclasts, together with collagen type I and high mineral content (69%) of which 99% is hydroxyapatite”<-Fibrocartilage may not be cartilage but it should should be capable of interestitial growth like growth plates.
“Enthesis fibrocartilage has been proposed to act as a barrier for cellular communication between tendon/ligament fibroblasts and osteocytes. This is due to the fact that fibrocartilage tissue is poorly vascularized and fibrochondrocytes do not express connexins and do not form gap junctions. Thus, intercellular communication needs to take place indirectly via cell–matrix interactions or soluble factors.”<-This will make getting stem cells from the enthesis into the bone slightly more challenging.
” the similarity in cellular and ECM composition between the fibrocartilage and the hyaline cartilage indicates an adaptation to compression. Consistently, mechanical loading plays an important role during enthesis maturation and contributes to the generation of a structural and cellular gradient at the insertion site. This gradation of structural composition turns into a gradation of mechanical properties that enables load transmission and reduces stress concentration”
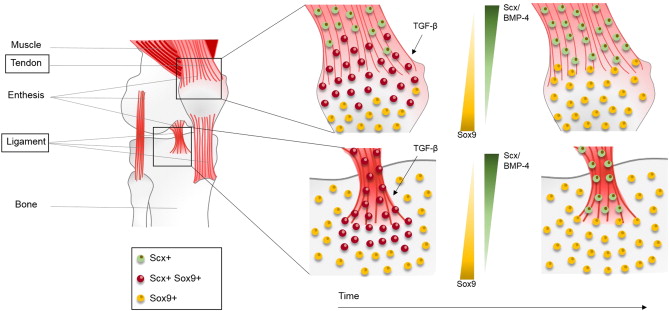
Enthesis cells express both Scx+ and Sox9+ progenitor cells. We have to encourage the enthesis to express more Sox9.
“During murine development, a progenitor cell population expressing both scleraxis (Scx) and Sox9 differentiates into Sox9 + chondrocytes or Scx + tenocytes/ligamentocytes to form the tendon/ligament-to-cartilage junction. Tenocytes are derived from both Scx +/Sox9 − and Scx +/Sox9 + progenitors, while ligamentocytes are derived exclusively from Scx +/Sox9 + progenitors[So ligaments always have chondrocyte differentiation capacity]. The closer the tendon is to the cartilaginous primordium, the more tenocytes arise from Scx +/Sox9 + progenitors. Similarly, enthesal chondrocytes located near the tendon/ligament-to-cartilage junction arise from Scx +/Sox9 + progenitors, while growth plate chondrocytes are derived from Scx −/Sox9 + progenitors. As development proceeds, the Scx +/Sox9 + progenitor cells upregulate either Scx expression, leading to cells of the tendon/ligament lineage, or Sox9 expression, leading to cells of the chondrocyte lineage. It has been proposed that TGF-β is necessary for Scx +/Sox9 + progenitor cell lineage specification, while the BMP-4/Scx axis determines its differentiation.”
“Fibrochondrocyte cells are expected to show a higher expression of collagen type II and lower expression of collagen type I when compared to fibroblasts.
Hypertrophic fibrochondrocytes synthetize higher levels of collagen type X.
Chondrocytes produce more collagen type II and less collagen type I than fibrochondrocytes.”
According to this paper, a crocodile may have a fibrocartilagenous growth plate which means that creating such a growth plate for humans is possible:
Crocodilian bone-tendon and bone-ligament interfaces
“We investigated bone-tendon (27 sites) and bone-ligament (12 sites) interfaces in six pairs of crocodile limbs and girdles under light microscopy. These crocodilian interfaces often included a direct, unmediated insertion in which the tendon or ligament fibers inserted directly into the bone itself without fibrocartilaginous mediation. This was quite different from the usual direct insertion known in mammals and lizards. Fibrocartilaginous tissue at the bone-tendon interface is generally believed to protect tendon fibers against shear stress. Other types of insertions were found in the crocodilian epiphyses, namely, hyaline cartilage and pseudofibrocartilaginous insertions. Notably, a thick periosteum/perichondrium and subchondral layer was involved at both interfaces. The thick periosteum/perichondrium seemed to form along the epiphyseal hyaline cartilage and might function in replacement of fibrocartilaginous tissues. Crocodilian thick periosterum/perichondrium would be expected to reinforce the limb and girdle bones — especially their epiphyses, in which secondary centers of ossification are absent. The subchondral layer — a kind of fibrocartila-ginous tissue — seemed to play the role of the growth plate in compensating for the absence of secondary centers of ossification. Therefore, we hypothesized that the crocodile-specific bone-tendon interfaces were the result of these specializations of bone development and growth. In crocodiles, the disadvantages of the single ossification center are effectively compensated for by specialized morphologies, including these interfaces. Specialized bone growth provides the crocodile with the largest body size of the recent reptiles and an extremely fast method of locomotion.”
Although according to the book Fins into Limbs: Evolution, Development, and Transformation avian growth plates become fibrocartilagenous when they are done growing.
The human tibial tubercle is formed by fibrous growth plates.
Here’s a grant about Scx and enthesis:
THE ROLE OF SCLERAXIS AND MECHANICAL LOADING ON ENTHESIS MATURATION
“The development of a functional attachment between tendon and bone (the “enthesis”) is critical for transmitting muscle forces to bone for joint motion. The formation of this attachment is driven by mechanical and biologic factors. Murine models of have demonstrated that removal of muscle load dramatically impairs the development of the enthesis. Furthermore, deletion of scleraxis (Scx), a transcription factor necessary for the development of force-transmitting tendons, leads to defects in enthesis formation. However, the mechanisms driving the mechanosensitivity of enthesis development are not known. Our global hypothesis is that Scx necessary for enthesis development and is driven by mechanical cues during{Maybe we can alter mechanical cues to alter enthesis development}. The first aim of this study is to determine the temporal influence of Scx on neonatal enthesis maturation using an inducible knockout murine model. Expression of Scx will be limited to development up to embryonic day 15.5, post-natal day 1, or post-natal day 7 and developmental adaptations and maturation of the enthesis will be assessed throughout juvenile development. It is expected that deletion of Scx during embryonic development will have a greater effect on enthesis maturation compared to post-natal deletion, and molecular and functional outcomes will be correlated with duration of Scx expression during maturation. These findings will establish the importance of Scx in enthesis development as well as determine its temporal role in post- natal musculoskeletal development. The second aim of this study is to determine the necessity of mechanical loading on Scx expression during development. Using in vivo and in vitro methods, we plan to mimic unloading and overloading, respectively, as well as assess the mechanosensitivity of Scx and its role in maturation and function of the enthesis. Using previously validated models of in vivo unloading and tissue-engineered methods for in vitro dynamic loading, we will track Scx-expressing cells as well as potential differentiation of ASCs into tenocytes. The effect of perturbed loading on the molecular and morphological response of Scx-expressing cells will be correlated with adaptations in structure and function using mechanical tests. These findings will determine the role of mechanical loading in Scx expression as well as the necessity of Scx in mechanically- induced tenogenesis. Lastly, the potency of Scx in load-induced tenogenesis following modulation with growth factors such as BMP-12 will be confirmed using BMP-12 siRNA. Findings from these studies will establish the importance of Scx on post-natal enthesis development, the mechanoresponsiveness of Scx in multi-tiered experimental models, and ultimately lead to new approaches for investigating musculoskeletal development and tissue engineered strategies for mechanosensitive orthopaedic tissues.”
Functional Attachment of Soft Tissues to Bone: Development, Healing, and Tissue Engineering

“Fibrous insertions typically occur over large areas, presumably to distribute forces and reduce stresses, and are characterized by perforating mineralized collagen fibers”
“The first zone consists of tendon/ligament, is populated by fibroblasts, and has mechanical properties similar to those of the tendon/ligament mid-substance. It is composed of linearly arrayed type I collagen fibers. The second zone consists of fibrocartilage, is populated by fibrochondrocytes, and is composed of types I, II, and III collagen and the proteoglycan aggrecan. The third zone is also populated by fibro-chondrocytes and is composed of mineralized fibrocartilage. Here, type II collagen predominates, and there are significant amounts of type X collagen as well as aggrecan. The fourth zone consists of bone proper, within which osteoblasts, osteocytes, and osteoclasts reside in a matrix of mineralized type I collagen.”
“the stratified tissue regions are compositionally distinct but structurally continuous.”<-thus anything that pulls on the tendon/ligament pulls on the bone.
“Enthesis development is initially driven by endochondral ossification: Cartilage mineralizes to form bone, and a fibrocartilaginous transition then develops at the interfaces between the bone and connective tissues.”
“All of the cell types found along the enthesis are mechanoresponsive; therefore, a role for mechanobiology during the development of this tissue is expected. Studies using genetically modified mice with muscular defects demonstrated that, although muscle loading was not required for initiation of enthesis formation, it was necessary for the subsequent growth and maturation of the enthesis”
“The reduction in muscle loading impaired mineral deposition and fibrocartilage formation and led to disorganized fiber distribution and inferior mechanical properties at the developing tendon enthesis.”
“mineralization at the developing enthesis is sensitive to mechanical loading.”
“when calcium phosphate cement was used to fill the tendon-to-bone junction in a rabbit ACL reconstruction model, the ceramic augmented bone tissue growth and organization”
“After the periosteal flap was sutured to the end of a torn rabbit infraspinatus tendon and then attached to bone, a fibrocartilage-like matrix developed after 12 weeks”