Update Nov 5,2014: Some people have commented that I am now monetizing the website. The negative reaction is a little surprising. The real reason I am putting everything in this book is because I am afraid of the possibility that all this knowledge which would be very helpful to humanity could be lost if something happened to me or something happened to the website. If hope the regular readers remember what happened to all that knowledge that was accumulated on the now dead GrowTallForum.com discussion boards. 5-6 years worth of information and ideas completely disappeared in 1 day after the admin decided to stop pushing forth with the endeavor.
Every time I get on an airplane and feel turbulence, I start to wonder what I have contributed to the world. How would I be remembered if I went down with the airplane? Did I make a positive change?
Maybe the book I plan to write is a way for me to leave something behind for the next generation of researchers, in case I suffer a horrible accident. This website has recently gone through at least 2 website technical problems, shutting down the website. I have come to realize that almost everything that is on the internet is just not on stable ground. At least if the information I will combine together is in PDF format and distributed to thousands of people around the world, the information would survive.
If the book was in real paperback or hardcover form, it could survive even the apocalypse, since the information and research I have gathered in the last 2 years will always be available.
I am currently working on writing a book that summarizes EVERYTHING that is on this website. All the important things will be added, and the extraneous information will not be. I will mention all the research and work that has been attempted in the last 15 years, from Sky, to Tyler, to Hakker, to Tim, and others who have tried to help out.
That is why I will not be posting on this website for at least another 4-6 months.
In addition, I have to focus on two other different businesses which are my primary sources of income.
Most of my time in the day is devoted to my companies and earning money, and keeping fit.
Most of my time at night will be devoted to finishing this book, as well as caring for my family members.
When the book is finally finished, probably in 2016, it will be released to the world, and this website will be transformed into something completely different.
I have decided to use the Natural Height Growth as one of the brand names under my primary umbrella corporation, and treat this website as a real asset. I will be coming back to make good on my promise to push the research much further.
This website will keep on going, and I will try to clean things up, and change dead links, and similar maintenance stuff now.
Anything new that comes out in the next few months will be from Tyler.
“Osteoporosis is a major public health issue that is expected to rise as the global population ages. Resveratrol (RES) is a plant polyphenol with various anti-aging properties. RES treatment of bone cells results in protective effects, but dose translation from in vitro studies to clinically relevant doses is limited since bioavailability is not taken into account. The aims of this review is to evaluate in vivo evidence for a role of RES supplementation in promoting bone health to reduced osteoporosis risk and potential mechanisms of action. Due to multiple actions on both osteoblasts and osteoclasts, RES has potential to attenuate bone loss resulting from different etiologies and pathologies. Several animal models have investigated the bone protective effects of RES supplementation. Ovariectomized rodent models of rapid bone loss due to estrogen-deficiency reported that RES supplementation improved bone mass and trabecular bone without stimulating other estrogen-sensitive tissues. RES supplementation prior to age-related bone loss was beneficial. The hindlimb unloaded rat model used to investigate bone loss due to mechanical unloading showed RES supplementation attenuated bone loss in old rats, but had inconsistent bone effects in mature rats. In growing rodents, RES increased longitudinal bone growth, but had no other effects on bone. In the absence of human clinical trials, evidence for a role of RES on bone heath relies on evidence generated by animal studies.”
“Resveratrol (RES) is a polyphenolic (3,4’,5-trihydroxystilbene) compound naturally present in red wine and a variety of plant foods such as grapes, cranberries, and nuts”
” human bone marrow-derived MSC with RES increased gene expression of the key osteogenic transcription factors, Runx2 and Osterix. RES was also demonstrated in vitro to act on various signal transduction pathways. RES activated the estrogen-mediated extracellular signal-regulated kinase 1/2 (ERK) signaling pathway regulating osteoblast differentiation and proliferation. RES activated AMP-activated protein kinase (AMPK) which regulates osteoblast differentiation and inhibits bone resorption by acting as a negative regulator of RANKL. RES augmented Wnt signaling which stimulated osteoblastogenesis and bone formation. Treating human bone marrow-derived MSC with RES promoted differentiation of MSC towards osteoblasts by up-regulating Runx2 gene expression through the activation of Sirt1. Also, activation of Sirt1 by RES was shown to promote binding to PPARγ which repressed MSC differentiation into adipocytes. Additionally, RES suppresses osteoclastogenesis by acting through Sirt1 to bind to RANK which inhibited binding to RANKL”<-many of these processes should impact longitudinal bone growth as well.
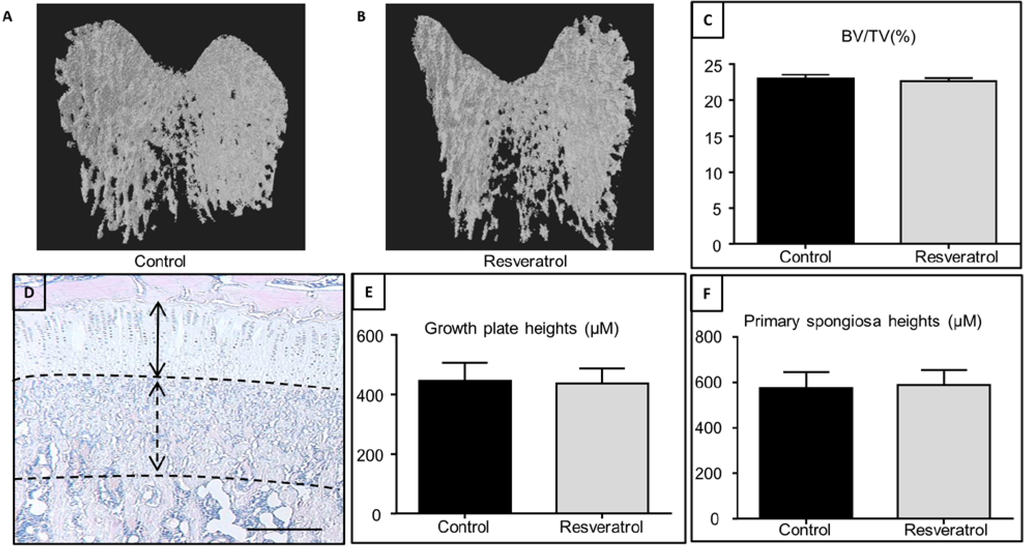
Resveratrol stimulated tibial and vertabral length in new zealand white rabbits that were 12 weeks old. 200mg per kg of bodyweight were given. Increased the amount of chondrocytes in the tibia and stimulated growth plate area while reducing fusion. Decreased vascularization indicated by lower VEGF and laminin levels.
“RES supplementation delayed growth plate fusion by suppressing the replacement of avascular cartilage with vascularized bone indicated by the down-regulated gene expression of vascular endothelial growth factor, a signaling molecule in vascularization, and laminin, a cartilage protein.”
In 6 month old Fisher-Brown Norway rats, it increased tibia length and width. Dosage was 12.5mg per kg of bodyweight.
“In vitro, RES treatment of chondrocytes obtained from an adult rat femur protected against the catabolic effect of pro-inflammatory cytokine, interleukin-1β”
“Osteoporosis is a major public health issue that is expected to rise as the global population ages. Resveratrol (RES) is a plant polyphenol with various anti-aging properties. RES treatment of bone cells results in protective effects, but dose translation from in vitro studies to clinically relevant doses is limited since bioavailability is not taken into account. The aims of this review is to evaluate in vivo evidence for a role of RES supplementation in promoting bone health to reduced osteoporosis risk and potential mechanisms of action. Due to multiple actions on both osteoblasts and osteoclasts, RES has potential to attenuate bone loss resulting from different etiologies and pathologies. Several animal models have investigated the bone protective effects of RES supplementation. Ovariectomized rodent models of rapid bone loss due to estrogen-deficiency reported that RES supplementation improved bone mass and trabecular bone without stimulating other estrogen-sensitive tissues. RES supplementation prior to age-related bone loss was beneficial. The hindlimb unloaded rat model used to investigate bone loss due to mechanical unloading showed RES supplementation attenuated bone loss in old rats, but had inconsistent bone effects in mature rats. In growing rodents, RES increased longitudinal bone growth, but had no other effects on bone. In the absence of human clinical trials, evidence for a role of RES on bone heath relies on evidence generated by animal studies. A better understanding of efficacy, safety, and molecular mechanisms of RES on bone will contribute to the determination of dietary recommendations and therapies to reduce osteoporosis.”
“eriosteal cortical bone formation coupled with endosteal cortical bone resorption regulates cross-sectional bone growth . To study bone growth, weanling female Sprague–Dawley rats were randomly assigned to a daily oral gavage of 0, 1, 4, 10, 40, 100 μg/d RES or estradiol (100 μg/d) dissolved in ethanol for a duration of 6 d. Despite estrogenic activity, RES had no significant effect on tibia cross-sectional area, medullary area, cortical bone area, periosteal bone formation rate or periosteal mineral apposition rat”
“In vitro, RES treatment of chondrocytes obtained from an adult rat femur protected against the catabolic effect of pro-inflammatory cytokine, interleukin-1B. To determine the effects of RES on longitudinal growth in vivo, a daily oral gavage of 200 mg/kg bwt RES [was given] to pubertal female New Zealand white rabbits until growth plate fusion occurred. After 16 weeks, rabbits provided RES supplementation had longer tibia and vertebrae, more chondrocytes, and increased growth plate area compared to control rabbits”
” Resveratrol, a natural polyphenol compound, may have the potential to promote bone formation and reduce bone resorption.”
“Resveratrol supplementation in the early ageing rats tended to decrease trabecular bone volume, Sirt1 gene expression and increased expression of adipogenesis-related genes in bone, all of which were statistically insignificant.”<- we want to increase sirt 1 levels.
“resveratrol supplementation does not significantly affect bone volume during the rapid growth phase but may potentially have negative effects on male skeleton during early ageing.”
In this study resveratrol did not impact growth plate height.
“Resveratrol has been identified as a potent activator of Sirtuin 1 (Sirt1), which is also known as nicotinamide adenine dinucleotide (NAD)-dependent deacetylase”
“resveratrol supplementation for 5 weeks during the rapid growth period in male rats had no significant effects on growth plate thickness, primary spongiosa heights and trabecular bone volume by the end of treatment, suggesting a lack of effect of resveratrol supplementation on the bone mass outcome in growing rats. These findings are in agreement with a previous study using an isoflavone-enriched diet containing soybean extract, daidzein, genistein, and equol in 6-week-old growing female pigs, which also found no significant changes in the growth plate, mineralization or osteoblast/osteoclast densities in long bones”
Resveratrol inhibits adipocyte differentiation and cellular senescence of human bone marrow stromal stem cells
“Resveratrol up-regulates osteogenic differentiation of hBMSCs, it may contribute to protection against bone loss. Resveratrol targets senescence, oxidative stress and up regulates endogenous protective anti-oxidant pathway in hBMSCs.”<-reversing senescence may keep things like the growth plates open for longer.
“Resveratrol (RSV), a plant-derived antioxidant mediating biological effects via sirtuin- related mechanisms”
“RSV reduced the levels of senescence-associated secretory phenotype (SASP), gene markers associated with senescence (P53, P16, and P21), intracellular ROS levels and increased gene expression of enzymes protecting cells from oxidative damage (HMOX1 and SOD3).”
Mimicking the Biochemical and Mechanical Extracellular Environment of the Endochondral Ossification Process to Enhance the In Vitro Mineralization Potential of Human Mesenchymal Stem Cells.
” Chondrogenesis and mechanical stimulation of the cartilage template are essential for bone formation through the endochondral ossification process in vivo. Recent studies have demonstrated that in vitro regeneration strategies that mimic these aspects separately, either chondrogenesis or mechanical stimulation, can promote mineralization to a certain extent both in vitro and in vivo. However, to date no study has sought to incorporate both the formation of the cartilage template and the application of mechanical stimulation simultaneously to induce osteogenesis. In this study, we test the hypothesis that mimicking both the biochemical and mechanical extracellular environment arising during endochondral ossification can enhance the in vitro mineralization potential of human mesenchymal stem cells (hMSCs). hMSC aggregates were cultured for 21 days under the following culture conditions; (1) Growth Medium – hydrostatic pressure (HP), (2) Chondrogenic Priming-HP, (3) Growth Medium + HP, and (4) Chondrogenic Priming +HP. Each group was then further cultured for another 21 days in the presence of osteogenic growth factors without HP. Biochemical (DNA, sulfate glycosaminoglycan, hydroxyproline, alkaline phosphatase activity, and calcium), histological (Alcian Blue and Alizarin Red), and immunohistological (Col I, II, and X, and BSP-2) analyses were conducted to investigate chondrogenic and osteogenic differentiation at various time points (14, 21, 35, and 42 days). Our results showed the application of HP-induced chondrogenesis similar to that of chondrogenic priming, but interestingly, there was a reduction in hypertrophy markers (collagen type X) by applying HP alone versus chondrogenic priming alone. Moreover, the results showed that both chondrogenic priming and HP in tandem during the priming period, followed by culture in osteogenic medium, accelerated the osteogenic potential of hMSCs.”
“application of hydrostatic pressure (0.1 MPa – 10 MPa) to human bone marrow stem cells aggregates or those seeded on a collagen or agarose scaffold can significantly enhance expression of chondrogenic markers (aggrecan, SOX‐9). The highest amount of matrix deposition was found when a pressure of 10 MPa was applied for a minimum of 5 days”
” the application of HP without any external growth factors resulted in reduced hypertrophy, whilst allowing for chondrogenesis, however once the HP was removed and the cells were exposed to osteogenic growth factors the cells began to produce hypertrophic markers but at a much slower rate than those exposed to chondrogenic growth factors alone”
“Pluripotent embryonic stem cells (ESCs) generate rostral paraxial mesoderm-like progeny in 5-6 days of differentiation induced by Wnt3a and Noggin (Nog). canonical Wnt signaling introduced either by forced expression of activated β-catenin, or the small-molecule inhibitor of Gsk3, CHIR99021, satisfied the need for Wnt3a signaling, and that the small-molecule inhibitor of BMP type I receptors, LDN193189, was able to replace Nog{so abnormal methods are able to replicate the bodies processes}. Mesodermal progeny generated using such small molecules were chondrogenic in vitro, and expressed trunk paraxial mesoderm markers such as Tcf15 and Meox1, and somite markers such as Uncx, but failed to express sclerotome markers such as Pax1. Induction of the osteochondrogenically committed sclerotome from somite requires sonic hedgehog and Nog. Consistently, Pax1 and Bapx1 expression was induced when the isolated paraxial mesodermal progeny were treated with SAG1 (a hedgehog receptor agonist) and LDN193189, then Sox9 expression was induced, leading to cartilaginous nodules and particles in the presence of BMP, indicative of chondrogenesis via sclerotome specification. By contrast, treatment with TGFβ also supported chondrogenesis and stimulated Sox9 expression, but failed to induce the expression of Pax1{Pax1 is upregulated by LSJL} and Bapx1. On ectopic transplantation to immunocompromised mice, the cartilage particles developed under either condition became similarly mineralized and formed pieces of bone with marrow. Thus, the use of small molecules led to the effective generation from ESCs of paraxial mesodermal progeny, and to their further differentiation in vitro through sclerotome specification into growth plate-like chondrocytes, a mechanism resembling in vivo somitic chondrogenesis that is not recapitulated with TGFβ. ”
“The osteochondro-progenitors that develop during embryogenesis are limb bud mesenchyme (derived from lateral plate mesoderm) responsible for limb bone and cartilage generation, sclerotome (from somite/rostral paraxial mesoderm) responsible for rib, vertebral joint, intervertebral disc and vertebral body formation, and ectomesenchyme (from cranial neural crest) responsible for craniofacial bone and cartilage generation.”
“The Flk1−Pdgfrα+ rostral paraxial mesoderm from ESCs consistently show chondrogenic activity in vitro ”
“SAG+LDN (PSL) stimulation during the first 6 days of micromass culture was crucial for inducing Pax1 and Bapx1 expression from the isolated E-cadherin−Flk1−Pdgfrα+ rostral paraxial mesoderm, as was the Shh+Nog stimulation”
“BMP signaling counteracts Shh+Nog and inhibits sclerotome induction”
Go to this page to see the original x-rays. Here’s a picture that shows that my right finger is longer than my left. However, what’s interesting is that the phalanx bones of the left index finger are actually longer than the right index finger. So, I turned to the metacarpal bone of the index fingers and it turns out that the right metacarpal of the index finger is longer than the left and a longer metacarpal would explain the increased finger length in the image in the second link below.
LSJL may have lengthened all the bones of my right finger but because my left finger was always longer it may have only reduced the discrepancy. If every single bone on the left side of body is longer than the right except for those that I performed LSJL on I submit that as proof of LSJL. A possible reason that all my left bones are longer than my right bones but all my right bones are thicker could be related to FGFR3 and CNP. My left side could have had less of the FGFR3 receptors or more of the CNP receptors.
I didn’t know that all my left sided bones were longer than my right until recently. I just assumed it was due to my scoliosis or it was just an illusion because my right side was thicker. So unfortunately, I did not account for it when designing to the experiment. But again, if every single bone on my hands is longer on my left than my right metacarpal which is where I performed LSJL then we can take that as evidence that LSJL works.
Here’s a video with my right finger before any loading.:
Here’s the left hand before video:
My right index finger metacarpal did grow longer but it would appear that my other bones did not. I say appear because it turns out that all my bones on the left side of my body are longer than the left. Thus, any growth may actually just be shown by decreasing the gap between the right and left finger bones. The right finger bones may have grown but that would not show up because the left finger bones were still longer. If every single bone has the version on the left side be longer, but the one bone that had LSJL on it can we say that LSJL induced growth?
Here are the metacarpals of my right and left index fingers:
The lines are very straight. I got pretty close to measuring out straight lines. I measured the right metacarpal as being 1383.0 pixels long. I got 1372.0 pixels long for the left metacarpal. The left metacarpal might extend a little below the line but I zoomed in so I could get things accurate. Right metacarpal is 0.8% longer. Now here’s thing about the right finger being 0.8% longer. All my left bones in my body are longer than the right bones but the right bones are thicker. Thus, the right metacarpal may have grown to catch up to the left metacarpal and then grown further. So actual growth may be larger than that.
I performed LSJL only on my right finger. Here’s an analysis of the other bones of the index finger:
The slope of the line may be off by about 0.11 degrees or so which is very minor. The left finger actually measures out longer.
Here’s an image of my proximal index fingers:This is a cut out of the proximal index fingers of the right and left finger bones from the x-ray from the link provided above. Click on the image as the image shows flipped for some reason. The right bone is the one labeled with the R. The measurements were done by adjusting the two bones until they were as long as possible. This insures that both bones were given the same advantages and the angle at which I put my finger is not a factor. Measurements were done by someone else.
The lines on the graph are slightly off the vertical lines sloping down at a 1.2 degree angle towards the right side of the computer screen(using the image if you click on the image not the inverted one here). This images slope favors the right side so the right side will appear longer if you don’t correct for the one degree.
Here’s another image:
This one slopes to the right only at about .80 degrees(measuring the straight line downwards it is .80 off from 90 degrees. This images slope favors the right as well but not nearly as much. The left bone is longer.
Here’s the cutout of the lateral view:
You can see here the the epiphysis of the right index finger are much larger but the left finger is longer overall.
How I’m changing my performance of LSJL now: Next step is to confirm that all my left handed bones are longer except for that metacarpal.
Now since I have the x-rays, I’m going to be performing LSJL on both my right and left index finger. Since I have x-ray images, I have those as a control using one of the fingers as a contralateral control is less important.
Previously, I was limiting my clamping based on my left side since my right side is stronger and thicker but since my left side is longer I’m not going to be clamping with whatever I can handle safely for my right side and whatever I can handle safely with the left side and not worrying about clamping them with the same intensity or amount of time. Since my right side is so much stronger it should be able to withstand more clamping. Thus, it was my goal with this to correct for the discrepancy between the right and left side in addition to increasing bone length overall.
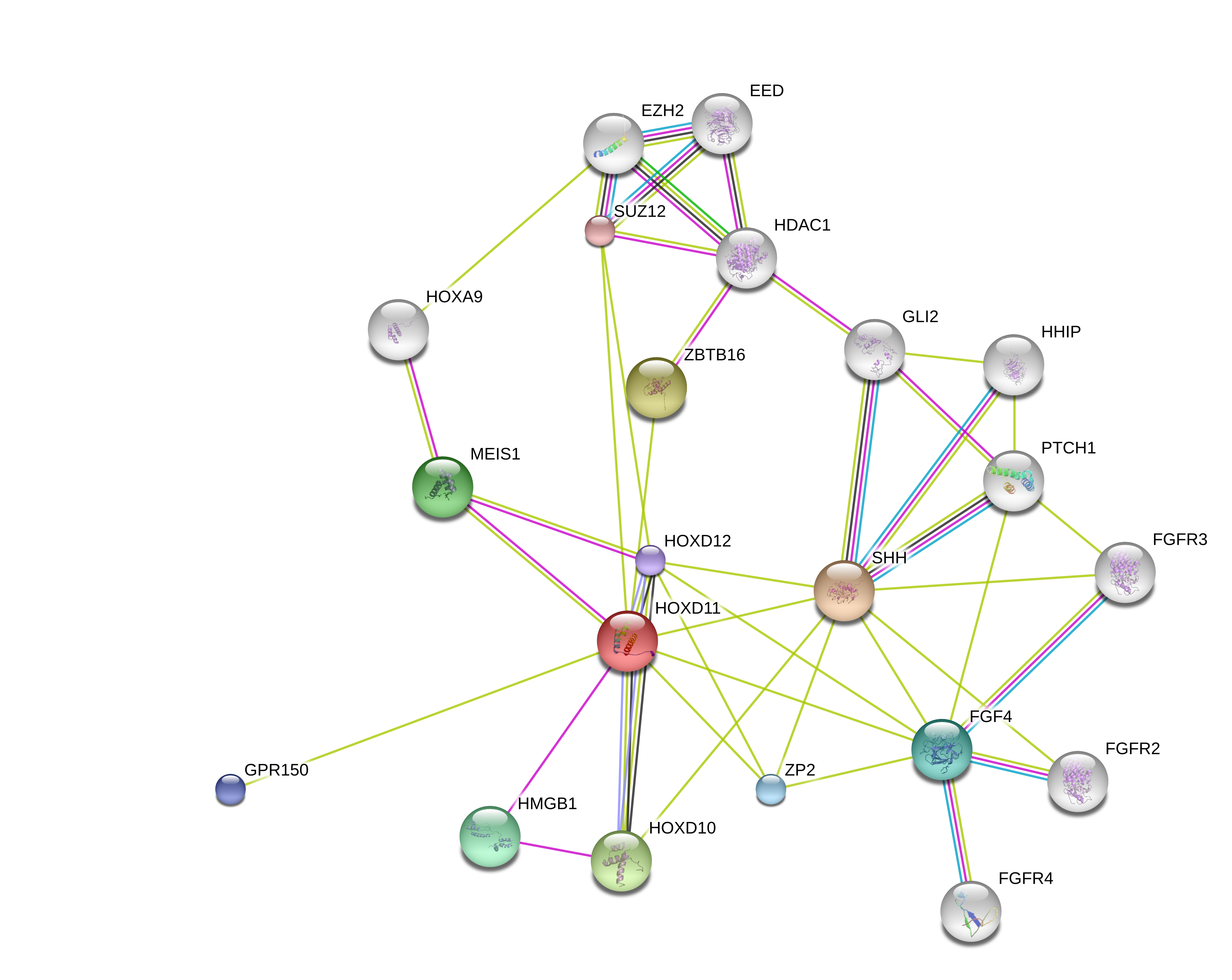
This study supports that altering gene expression of Hoxa11 and Hoxd11 may be a way to grow taller as those genes can turn short bones into long bones. Unfortunately, the genes that are known to interact with these genes are small in number and not known to be manipulatible by exercise or nutrition. So it is more likely that in the future the Hoxa11 and Hoxd11 genes may contribute to height growth by manipulating the expression of these genes via an adenovirus.
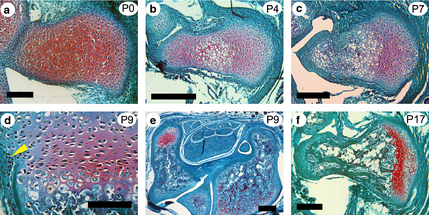
“The human pisiform is a small, nodular, although functionally significant, bone of the wrist. In most other mammals, including apes and Australopithecus afarensis, pisiforms are elongate. the typical mammalian pisiform forms from two ossification centers. We hypothesize that: (i) the presence of a secondary ossification center in mammalian pisiforms indicates the existence of a growth plate; and (ii) human pisiform reduction results from growth plate loss. We surveyed African ape pisiform ossification and confirmed the presence of a late-forming secondary ossification center in chimpanzees and gorillas. Identification of the initial ossification center occurs substantially earlier in apes relative to humans, raising questions concerning the homology of the human pisiform and the two mammalian ossification centers. Second, we conducted histological and immunohistochemical analyses of pisiform ossification in mice. We confirm the presence of two ossification centers separated by organized columnar and hypertrophic chondrocyte zones. Flattened chondrocytes were highly mitotic, indicating the presence of a growth plate. Hox genes have been proposed to play a fundamental role in growth plate patterning. The existence of a pisiform growth plate presents an interesting test case for the association between Hox expression and growth plate formation, and could explain the severe effects on the pisiform observed in Hoxa11 and Hoxd11 knockout mice. Hoxd11 is expressed adjacent to the pisiform in late-stage embryonic mouse limbs supporting a role for Hox genes in growth plate specification.”
“Compared with humans, the pisiform of most other mammals, including primates, is substantially enlarged and elongated”
Here’s a good visual of growth plate formation:
“a) At birth (P0) the pisiform largely consists of undifferentiated hyaline cartilage. Note the future articular surfaces adjacent to the triquetral (right) and the transitional region near the insertion of the FCU (left). Each of these is distinct from the fibrous periosteal layers that surround the future pisiform shaft. (b) At P4 the cartilage has undergone differentiation to flattened columnar and hypertrophic chondrocytes. It is the calcified hypertrophic matrix that is staining red in Fig. 3(a). (c) By P7 the primary center of ossification begins to be replaced by bone. A broad region of flattened columnar and hypertrophic chondrocytes is preserved at the palmar end (right). (d) At P9 the preserved strip of cartilage displays all of the hallmarks of a growth plate: organized columnar and hypertrophic zones and a perichondrial ring (yellow arrowhead) adjacent to the bone collar. (e) A transverse section through the carpal tunnel demonstrates the unique ossification of the pisiform (left). Note the preserved region of red stained cartilage at the palmar end. In contrast the scapholunate (right) has ossified as a single primary center extending into the projecting tubercle. (f) At P17 the growth plate appears to be losing its activity, as there is no longer an identifiable hypertrophic zone underlying the columnar chondrocytes.”<-so if via LSJL we can induce regions of hyaline cartilage they could potentially become growth plates.
“Full deletion of Hoxa11 or Hoxd11 results in a highly penetrant phenotype with shortened pisiforms that often fuse to the triquetral (ulnare) or less commonly to the scapholunate and triquetral ”
” ‘no Hoxd land’ for short bone morphology”<-Maybe we could upregulate Hoxd in short bones to make them become long bones?
“In short bones and epiphyses, the initial process of chondrocyte differentiation is similar. However, expanded cartilaginous growth plates or active perichondrial rings do not form. Instead, the periphery of these regions largely consists of a narrow three to four cell layer of round chondrocytes that anticipate the future articular zone. In each of these respects (organized chondrocyte zones, active perichondrial ring and deposition of the bone collar), the pisiform is more similar to long bones”<-The organized chondrocyte zones, active perichondrial ring and deposition of the bone collar could be related to Hoxd.
“mice with reduced Hoxa11/Hoxd11 expression display decreased proliferation within mesenchymal condensations and dramatic shortening of the radius and ulna such that they also resemble short bones”
If we can induce Hoxd11 expression in short bones we can make those bones longer and thus ourselves taller.
Looking at the String Embl gene interaction reveals a problem in that very few genes directly interact with Hoxd11. Although LSJL upregulates the related gene Hoxd10: